第3章高考强化
一、刷真题
1.下列关于“ $ \mathrm{D}\mathrm{N}\mathrm{A} $ 粗提取与鉴定”实验的叙述,错误的是( )
A.实验中如果将研磨液更换为蒸馏水, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 提取的效率会降低
B.利用 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 和蛋白质在酒精中的溶解度差异,可初步分离 $ \mathrm{D}\mathrm{N}\mathrm{A} $
C. $ \mathrm{D}\mathrm{N}\mathrm{A} $ 在不同浓度 $ \mathrm{N}\mathrm{a}\mathrm{C}\mathrm{l} $ 溶液中溶解度不同,该原理可用于纯化 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 粗提物
D.将溶解的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 粗提物与二苯胺试剂反应,可检测溶液中是否含有蛋白质杂质
答案:D
解析:研磨液中的 $ \mathrm{S}\mathrm{D}\mathrm{S} $ 可使蛋白质变性,与 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分离; $ \mathrm{E}\mathrm{D}\mathrm{T}\mathrm{A} $ 是 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 酶抑制剂,可防止细胞破碎后 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 酶降解 $ \mathrm{D}\mathrm{N}\mathrm{A} $ ,故实验中如果将研磨液更换为蒸馏水, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 提取的效率会降低, $ \mathrm{A} $ 正确。 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 不溶于酒精,但某些蛋白质溶于酒精,利用这一原理可初步分离 $ \mathrm{D}\mathrm{N}\mathrm{A} $ , $ \mathrm{B} $ 正确。 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 在不同浓度 $ \mathrm{N}\mathrm{a}\mathrm{C}\mathrm{l} $ 溶液中溶解度不同,该原理可用于纯化 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 粗提物, $ \mathrm{C} $ 正确。在一定温度下, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 遇二苯胺试剂会呈现蓝色,因此二苯胺试剂可作为鉴定 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 的试剂,但不可检测溶液中是否含有蛋白质杂质, $ \mathrm{D} $ 错误。
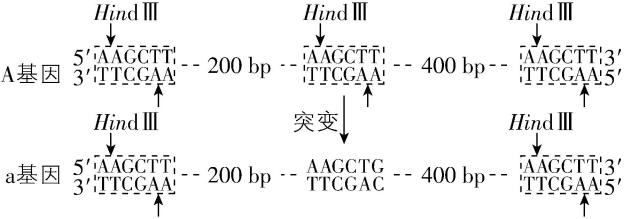
2. 2.(多选题)图示人体正常基因A突变为致病基因 $ \mathrm{a} $ 及 $ Hin\mathrm{d}Ⅲ $ 切割位点。 $ Alu $ Ⅰ限制酶识别序列及切割位点为 ,下列相关叙述正确的有( )
,下列相关叙述正确的有( )
 (多选)
(多选)
A.基因A突变为 $ \mathrm{a} $ 是一种碱基增添的突变
B.用两种限制酶分别酶切A基因后,形成的末端类型不同
C.用两种限制酶分别酶切 $ \mathrm{a} $ 基因后,产生的片段大小一致
D.产前诊断时,该致病基因可选用 $ Hin\mathrm{d}Ⅲ $ 限制酶开展酶切鉴定
答案:BD
解析:分析题图可知,基因 $ \mathrm{A} $ 突变为致病基因 $ \mathrm{a} $ 是 $ \mathrm{T}—\mathrm{A} $ 被 $ \mathrm{G}—\mathrm{C} $ 替换造成的, $ \mathrm{A} $ 错误;用限制酶 $ Hin\mathrm{d}Ⅲ $ 酶切 $ \mathrm{A} $ 基因后会形成黏性末端,用限制酶 $ Alu $ Ⅰ酶切 $ \mathrm{A} $ 基因后,形成的是平末端, $ \mathrm{B} $ 正确。
3.质粒 $ \mathrm{K} $ 中含有 $ \beta - $ 半乳糖苷酶基因,将该质粒导入大肠杆菌细胞后,其编码的酶可分解 $ \mathrm{X}-\mathrm{g}\mathrm{a}\mathrm{l} $ ,产生蓝色物质,进而形成蓝色菌落,如图所示。科研小组以该质粒作为载体,采用基因工程技术实现人源干扰素基因在大肠杆菌中的高效表达。下列叙述错误的是( )
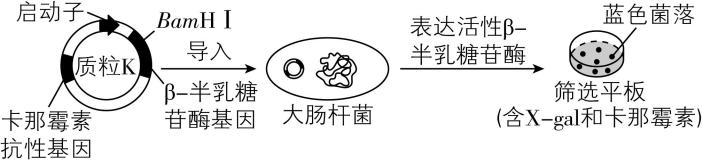
A.使用氯化钙处理大肠杆菌以提高转化效率,可增加筛选平板上白色和蓝色菌落数
B.如果筛选平板中仅含卡那霉素,生长出的白色菌落不可判定为含目的基因的菌株
C.因质粒 $ \mathrm{K} $ 中含两个标记基因,筛选平板中长出的白色菌落即为表达目标蛋白的菌株
D.若筛选平板中蓝色菌落偏多,原因可能是质粒 $ \mathrm{K} $ 经酶切后自身环化并导入了大肠杆菌
答案:C
解析:提高大肠杆菌转化效率的常用方法是使用氯化钙(或 $ {\mathrm{C}\mathrm{a}}^{2+} $ )处理细胞,使其处于一种能吸收周围环境中 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的生理状态,可增加筛选平板上白色和蓝色菌落数, $ \mathrm{A} $ 正确;由思路导引可知,若筛选平板中仅含卡那霉素,目的基因成功插入的重组质粒导入的大肠杆菌以及目的基因未插入成功的质粒 $ \mathrm{K} $ 导入的大肠杆菌在平板上均为白色菌落,故无法判断哪个是含目的基因的菌株, $ \mathrm{B} $ 正确;因质粒 $ \mathrm{K} $ 中含两个标记基因,筛选平板中长出的白色菌落应为含目的基因的菌株,能否表达相应蛋白,需进一步检测, $ \mathrm{C} $ 错误;如果筛选平板上蓝色菌落明显偏多,说明大量没有插入目的基因的质粒 $ \mathrm{K} $ 导入了大肠杆菌,推测原因可能是酶切后的载体发生自身环化并导入了大肠杆菌, $ \mathrm{D} $ 正确。
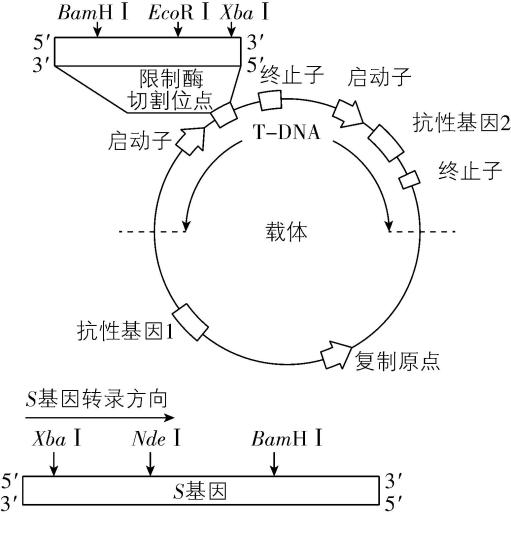
4.马铃薯作为重要农作物,提高其冷耐受性可拓展优质马铃薯的种植区域。我国科研人员发现,野生马铃薯中 $ S $ 基因的表达与其冷耐受性调控有关,将该基因导入栽培马铃薯中可显著增强其抗寒能力。回答下列问题。
限制酶 | 识别序列及切割位点 |
$ Bgl $ Ⅱ | 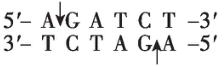
|
$ Nde $ Ⅰ | 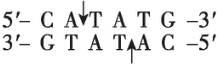
|
$ Xho $ Ⅰ | 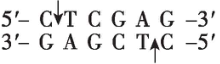
|
$ EeoR $ Ⅰ | 
|
$ BamH $ Ⅰ | 
|
$ Sal $ Ⅰ | 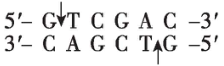
|
$ Xba $ Ⅰ | 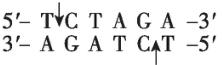
|
(1) $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增目的基因时,需要模板 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 、引物、 、含 $ {\mathrm{M}\mathrm{g}}^{2+} $ 的缓冲液和耐高温的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 聚合酶。 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 聚合酶在 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 的 步骤中起作用。
(2) 图中标识了载体和 $ S $ 基因中限制酶的切割位点。为将 $ S $ 基因正确插入载体, $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增 $ S $ 基因时需在引物的 (填“ $ 5\prime $ 端”或“ $ 3\prime $ 端”)添加限制酶识别序列,结合上表分析,上游引物应添加的碱基序列是 $ 5\prime - $ $ -3\prime $ ,切割载体时应选用的两种限制酶是 。 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增产物和载体分别被限制酶切割后,经纯化和连接,获得含 $ S $ 基因的表达载体并导入农杆菌。
(3) 用携带 $ S $ 基因的农杆菌侵染栽培马铃薯愈伤组织时,基因表达载体中 $ \mathrm{T}-\mathrm{D}\mathrm{N}\mathrm{A} $ 进入愈伤组织细胞,将 $ S $ 基因整合到 ,抗性基因 可用于筛选成功转化的愈伤组织。该愈伤组织经 形成芽、根,继续培育可获得抗寒能力显著增强的马铃薯植株。
答案:(1) 脱氧核苷酸;延伸
(2) $ 5\prime $ 端; $ \mathrm{A}\mathrm{G}\mathrm{A}\mathrm{T}\mathrm{C}\mathrm{T} $ ; $ Bam\mathrm{H}Ⅰ $ 与 $ Eco\mathrm{R}Ⅰ $
(3) 染色体 $ \mathrm{D}\mathrm{N}\mathrm{A} $ ;2;再分化
解析:(1) $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增目的基因时,需要模板 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 、引物、(4种)脱氧核苷酸、含 $ {\mathrm{M}\mathrm{g}}^{2+} $ 的缓冲液和耐高温的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 聚合酶。 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 过程一般可以分为变性、复性、延伸, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 聚合酶在延伸步骤中起作用,即 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 聚合酶将脱氧核苷酸加到引物的 $ 3\prime $ 端进行子链的延伸。
(2) 为将 $ S $ 基因正确插入载体, $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增 $ S $ 基因时需要在引物的 $ 5\prime $ 端添加相应的限制酶识别序列。为保证目的基因的正确插入,应使用双酶切;载体中含有 $ Bam\mathrm{H}Ⅰ $ 、 $ Eco\mathrm{R}Ⅰ $ 和 $ Xba $ Ⅰ的酶切位点, $ S $ 基因上含有 $ Xba $ Ⅰ、 $ Nde $ Ⅰ、 $ Bam\mathrm{H}Ⅰ $ 的酶切位点,为保证 $ S $ 基因结构的完整性,不能在 $ S $ 基因两端添加 $ Xba $ Ⅰ、 $ Nde $ Ⅰ、 $ Bam\mathrm{H}Ⅰ $ 的识别序列;分析表格数据, $ Bgl $ Ⅱ切割产生的黏性末端和 $ Bam\mathrm{H}Ⅰ $ 切割产生的黏性末端相同,结合 $ S $ 基因的转录方向和载体上启动子的转录方向可知,应在 $ S $ 基因上游添加 $ Bgl $ Ⅱ的识别序列,即 $ 5\prime -\mathrm{A}\mathrm{G}\mathrm{A}\mathrm{T}\mathrm{C}\mathrm{T}-3\prime $ ,下游添加 $ Eco\mathrm{R}Ⅰ $ 的识别序列。因此,切割载体应选用的两种限制酶为 $ Bam\mathrm{H}Ⅰ $ 和 $ Eco\mathrm{R}Ⅰ $ 。
(3) 用携带 $ S $ 基因的农杆菌侵染栽培马铃薯的愈伤组织时,农杆菌能将基因表达载体中的 $ \mathrm{T}-\mathrm{D}\mathrm{N}\mathrm{A} $ 转移到愈伤组织细胞的染色体 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 上。分析载体, $ \mathrm{T}-\mathrm{D}\mathrm{N}\mathrm{A} $ 中含有抗性基因2,若 $ \mathrm{T}-\mathrm{D}\mathrm{N}\mathrm{A} $ 成功整合到被侵染细胞的染色体 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 上,则该细胞具有抗性基因2对应的抗性,因此,抗性基因2可用于筛选成功转化的愈伤组织。该愈伤组织经再分化形成芽、根,继续培育可获得抗寒能力显著增加的马铃薯植株。
5.稻瘟病是一种真菌病害,水稻叶片某些内生放线菌对该致病菌有抑制作用。科研小组分离筛选出内生放线菌,并开展了相关研究。回答下列问题。
(1) 采集有病斑的水稻叶片,经表面消毒、研磨处理,制备研磨液。此后,采用 (填方法)将研磨液接种于不同的选择培养基,分别置于不同温度下培养,目的是 。
(2) 经筛选获得一株内生放线菌,该菌株高效合成铁载体小分子,能辅助内生放线菌吸收铁离子。 $ R $ 基因是合成铁载体的关键基因之一。科研小组构建 $ R $ 基因敲除株,探究铁载体的功能。主要步骤如下:首先克隆 $ R $ 基因的上游片段 $ R-U $ 和下游片段 $ R-D $ ;然后构建重组质粒;最后利用重组质粒和内生放线菌 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 片段中同源区段可发生交换的原理,对目标基因进行敲除。如图1所示。
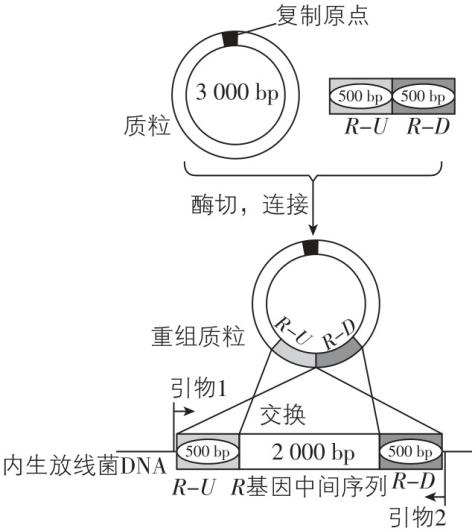
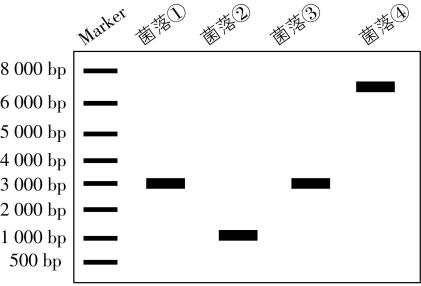
图1 图2
说明:用图1中的引物1和2进行 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增
采用 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 技术鉴定 $ R $ 基因的敲除结果。 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 通过变性、复性和延伸三步,反复循环,可实现基因片段的 。 $ R $ 基因敲除过程中,可发生多种形式的同源区段交换, $ \mathrm{P}\mathrm{C}\mathrm{R} $ 检测结果如图2所示,其中 $ R $ 基因敲除株为菌落 (填序号),出现菌落④的可能原因是 。
(3) 内生放线菌和稻瘟病致病菌的生长均需要铁元素。科研小组推测该内生放线菌通过对铁离子的竞争性利用,从而抑制稻瘟病致病菌生长。设计实验验证该推测,简要写出实验思路。
答案:(1) 稀释涂布平板法;探究内生放线菌最适的营养条件和温度
(2) 快速扩增;②;只有 $ R-U $ 或 $ R-D $ 一处发生了同源重组
(3) 将野生型内生放线菌、 $ R $ 基因敲除内生放线菌分别与稻瘟病致病菌混合培养在含铁培养液中,统计培养液中铁离子含量及两种菌的数量变化。
解析:(1) 可采用稀释涂布平板法将研磨液接种于不同的选择培养基,并分别置于不同温度下培养的目的是探究内生放线菌最适的营养条件和温度。
(2) $ \mathrm{P}\mathrm{C}\mathrm{R} $ 的实质是体外的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制,所以 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 可以实现基因片段的快速扩增。由图1可知,未发生同源重组前,在引物1、2的作用下可扩增出的片段为 $ 3000\mathrm{b}\mathrm{p} $ ,故菌落①③未发生同源重组。若 $ R-U $ 和 $ R-D $ 都发生同源重组,则会减少 $ R $ 基因中间的 $ 2000\mathrm{b}\mathrm{p} $ 的序列,从而使扩增的片段减小到 $ 1000\mathrm{b}\mathrm{p} $ ,故菌落②为发生同源重组后的 $ R $ 基因敲除株。根据题干信息, $ R $ 基因敲除过程中可发生多种形式的同源区段交换,菌落④的大小约为 $ 7000\mathrm{b}\mathrm{p} $ ,则可以推测在同源重组过程中,只发生了 $ R-U $ 或 $ R-D $ 一处同源重组,导致整个质粒片段接入。
(3) 若要验证内生放线菌通过与稻瘟病致病菌竞争性利用铁离子抑制稻瘟病致病菌的生长,则实验的自变量应为内生放线菌是否能利用培养液中的铁离子,即 $ R $ 基因的有无,因变量应该为稻瘟病致病菌的生长情况,最后可观测两种菌的数量和培养液中的铁离子含量变化情况来验证实验结论。所以实验可分为两组,甲组:将适量的含 $ R $ 基因的内生放线菌和稻瘟病致病菌混合培养在含铁的培养液中;乙组:将等量的 $ R $ 基因敲除内生放线菌与稻瘟病致病菌混合培养在相同的含铁培养液中;将两组培养液放在适宜的条件下,间隔合适时间记录每组培养液中铁离子含量变化和两种菌的数量变化。
6.香树脂醇具有抗炎等功效,从植物中提取难度大、产率低。通过在酵母菌中表达外源香树脂醇合酶基因 $ N $ ,可高效生产香树脂醇。回答下列问题。
(1) 可从 中查询基因 $ N $ 的编码序列,设计特定引物。如图1所示, $ \mathrm{a} $ 链为转录模板链,为保证基因 $ N $ 与质粒 $ pYL $ 正确连接,需在引物1和引物2的 $ 5\prime $ 端分别引入 和 限制酶识别序列。 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增基因 $ N $ ,特异性酶切后,利用 连接 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 片段,构建重组质粒,大小约 $ 9.5\mathrm{k}\mathrm{b}(\mathrm{k}\mathrm{b} $ 为千碱基对 $ ) $ ,假设构建重组质粒前后,质粒 $ pYL $ 对应部分大小基本不变。
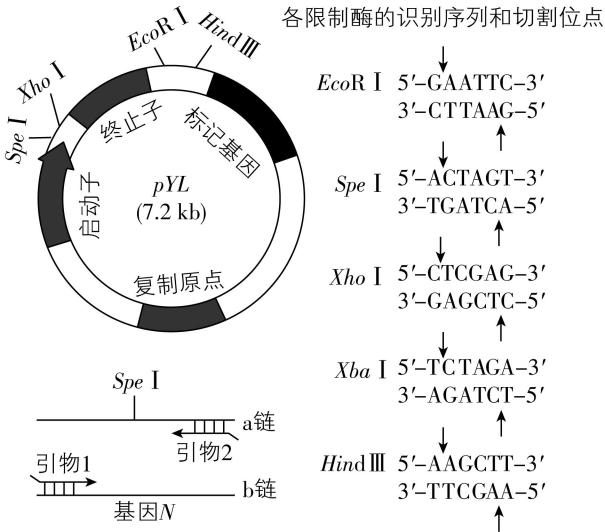
图1
(2) 进一步筛选构建的质粒,以 $ 1~4 $ 号菌株中提取的质粒为模板,使用引物1和引物2进行 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增,电泳 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 产物,结果如图2。在第5组的 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 反应中,使用无菌水代替实验组的模板 $ \mathrm{D}\mathrm{N}\mathrm{A} $ ,目的是检验 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 反应中是否有 的污染。初步判断实验组 (从“ $ 1\sim 4 $ ”中选填)的质粒中成功插入了基因 $ N $ ,理由是 。
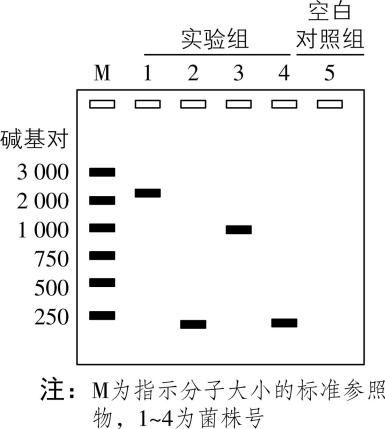
图2
(3) 为提高香树脂醇合酶催化效率,将编码第240位脯氨酸或第243位苯丙氨酸的碱基序列替换为编码丙氨酸的碱基序列,丙氨酸的密码子有 $ \mathrm{G}\mathrm{C}\mathrm{A} $ 等。 $ \mathrm{a} $ 是诱变第240位脯氨酸编码序列的引物( $ \mathrm{G}\mathrm{C}\mathrm{A} $ 为诱变序列), $ \mathrm{b} $ 、 $ \mathrm{c} $ 、 $ \mathrm{d} $ 其中一条是诱变第243位苯丙氨酸的引物,其配对模板与 $ \mathrm{a} $ 的配对模板相同。据此分析,丙氨酸的密码子除 $ \mathrm{G}\mathrm{C}\mathrm{A} $ 外,还有 。
$ \mathrm{a}:5\prime -\cdots \mathrm{G}\mathrm{C}\mathrm{A}/\mathrm{C}\mathrm{C}\mathrm{C}/\mathrm{G}\mathrm{A}\mathrm{G}/\mathrm{T}\mathrm{T}\mathrm{C}/\mathrm{T}\mathrm{G}\mathrm{G}/\mathrm{C}\mathrm{T}\mathrm{G}/\mathrm{T}\mathrm{T}\mathrm{T}/\mathrm{C}\mathrm{C}\mathrm{C}/\mathrm{T}\mathrm{C}\mathrm{T}/\mathrm{T}\mathrm{T}\mathrm{C}/\mathrm{T}\mathrm{T}\mathrm{C}\cdots -3\prime $
$ \mathrm{b}:5\prime -\cdots \mathrm{G}\mathrm{A}\mathrm{A}/\mathrm{C}\mathrm{T}\mathrm{G}/\mathrm{T}\mathrm{G}\mathrm{G}/\mathrm{G}\mathrm{A}\mathrm{C}/\mathrm{A}\mathrm{C}\mathrm{C}/\mathrm{C}\mathrm{T}\mathrm{G}/\mathrm{A}\mathrm{A}\mathrm{C}/\mathrm{T}\mathrm{A}\mathrm{C}/\mathrm{T}\mathrm{T}\mathrm{C}/\mathrm{T}\mathrm{C}\mathrm{T}/\mathrm{G}\mathrm{A}\mathrm{G}\cdots -3\prime $
$ \mathrm{c}:5\prime -\cdots \mathrm{G}\mathrm{C}\mathrm{C}/\mathrm{T}\mathrm{G}\mathrm{G}/\mathrm{C}\mathrm{T}\mathrm{G}/\mathrm{T}\mathrm{T}\mathrm{T}/\mathrm{C}\mathrm{C}\mathrm{C}/\mathrm{T}\mathrm{C}\mathrm{T}/\mathrm{T}\mathrm{T}\mathrm{C}/\mathrm{T}\mathrm{T}\mathrm{C}/\mathrm{C}\mathrm{C}\mathrm{C}/\mathrm{T}\mathrm{A}\mathrm{C}/\mathrm{C}\mathrm{A}\mathrm{C}\cdots -3\prime $
$ \mathrm{d}:5\prime -\cdots \mathrm{G}\mathrm{A}\mathrm{T}/\mathrm{A}\mathrm{A}\mathrm{T}/\mathrm{A}\mathrm{A}\mathrm{G}/\mathrm{A}\mathrm{T}\mathrm{C}/\mathrm{C}\mathrm{G}\mathrm{A}/\mathrm{G}\mathrm{A}\mathrm{G}/\mathrm{A}\mathrm{A}\mathrm{G}/\mathrm{G}\mathrm{C}\mathrm{C}/\mathrm{A}\mathrm{T}\mathrm{G}/\mathrm{C}\mathrm{G}\mathrm{A}/\mathrm{A}\mathrm{A}\mathrm{G}\cdots -3\prime $
(4) 进一步检测转基因酵母菌发酵得到的 含量并进行比较,可以选出最优的香树脂醇合酶基因的改造方案。
答案:(1) 基因序列数据库(或 $ \mathrm{G}\mathrm{e}\mathrm{n}\mathrm{B}\mathrm{a}\mathrm{n}\mathrm{k} $ 或 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 序列数据库); $ Xho $ Ⅰ; $ Xba $ Ⅰ; $ \mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶
(2) 外源 $ \mathrm{D}\mathrm{N}\mathrm{A} $ (或外源基因、基因 $ N $ );1;基因 $ N $ 大小为重组质粒大小和质粒 $ pYL $ 大小的差值,约为 $ 2.3\mathrm{k}\mathrm{b} $ ,对应实验组1的电泳条带(或实验组1电泳条带大小加质粒 $ pYL $ 大小约等于重组质粒大小)
(3) $ \mathrm{G}\mathrm{C}\mathrm{C} $
(4) 香树脂醇
解析:(1) 可从基因序列数据库中查询基因 $ N $ 的编码序列,设计特定引物。由思路导引可知,为保证基因 $ N $ 与质粒 $ pYL $ 正确连接,需在引物1和引物2的 $ 5\prime $ 端分别引入 $ Xho $ Ⅰ和 $ Xba $ Ⅰ限制酶识别序列。 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增基因 $ N $ ,经特异性酶切后,利用 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶连接 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 片段,构建重组质粒。
(2) 在第5组的 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 反应中,使用无菌水代替实验组的模板 $ \mathrm{D}\mathrm{N}\mathrm{A} $ ,目的是检验 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 反应中是否有外源 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 的污染。质粒 $ pYL $ 大小为 $ 7.2\mathrm{k}\mathrm{b} $ ,重组质粒大小约 $ 9.5\mathrm{k}\mathrm{b} $ ,假设构建重组质粒前后,质粒 $ pYL $ 对应部分大小基本不变,基因 $ N $ 大小为二者的差值,约为 $ 2.3\mathrm{k}\mathrm{b}(2\sim 2.3\mathrm{k}\mathrm{b}) $ ,结合图2电泳结果可知,实验组1的电泳条带大小约为 $ 2300\mathrm{b}\mathrm{p} $ ,说明实验组1的质粒中成功插入了基因 $ N $ 。
(3) $ \mathrm{a} $ 是诱变第240位脯氨酸编码序列替换为丙氨酸编码序列的引物( $ \mathrm{G}\mathrm{C}\mathrm{A} $ 为诱变序列,与丙氨酸的密码子序列相同), $ \mathrm{b} $ 、 $ \mathrm{c} $ 、 $ \mathrm{d} $ 其中一条是诱变第243位苯丙氨酸编码序列替换为丙氨酸编码序列的引物,其配对模板与 $ \mathrm{a} $ 的配对模板相同,据此分析,除编码第243位苯丙氨酸的碱基序列 $ \mathrm{T}\mathrm{T}\mathrm{C} $ 替换为诱变序列,与丙氨酸的密码子序列相同,其后的编码序列应该与 $ \mathrm{a} $ 相同,为 $ \mathrm{T}\mathrm{G}\mathrm{G}/\mathrm{C}\mathrm{T}\mathrm{G}/\mathrm{T}\mathrm{T}\mathrm{T}\cdots \cdots $ ,所以该引物是 $ \mathrm{c} $ ,其中 $ \mathrm{G}\mathrm{C}\mathrm{C} $ 为第243位诱变序列,故丙氨酸的密码子还可以是 $ \mathrm{G}\mathrm{C}\mathrm{C} $ 。
(4) 本题旨在通过在酵母菌中表达外源香树脂醇合酶基因 $ N $ 并进行改造,提高香树脂醇合酶的催化效率,高效生产香树脂醇,所以检测转基因酵母菌发酵产物香树脂醇的含量并进行比较,可以选出香树脂醇合酶催化效率较高的香树脂醇合酶基因改造方案。
7.卡拉胶是一类源于海洋红藻的大分子多糖,可被某些细菌降解为具有多种应用前景的卡拉胶寡糖。某研究小组拟筛选具有高活性卡拉胶酶 $ (\mathrm{C}\mathrm{G}) $ 的菌种用于生产卡拉胶寡糖。回答下列问题:
(1) 选择海藻和海泥作为样本筛选卡拉胶降解菌的原因是 。将培养后的菌液混匀并充分 ,再接种至微孔板中,经培养和筛选获得了 $ \mathrm{C}\mathrm{G} $ 活性最高的菌种。
(2) 为构建携带 $ cg(\mathrm{C}\mathrm{G} $ 的编码基因 $ ) $ 的大肠杆菌表达载体(如图),对 $ cg $ 的 $ \mathrm{P}\mathrm{C}\mathrm{R} $ 扩增产物和质粒进行双酶切,随后用 $ E.coli\mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶连接。为保证连接准确性和效率, $ cg $ 转录模板链的 $ 5\prime $ 端最好含有 酶切位点。另有两组同学选用了各不相同的双酶切组合和 $ \mathrm{T}4\mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶重复上述实验,获得的部分重组质粒分子大小符合预期,但均无法使用各自构建表达载体的双酶切组合进行切割,其原因是 。
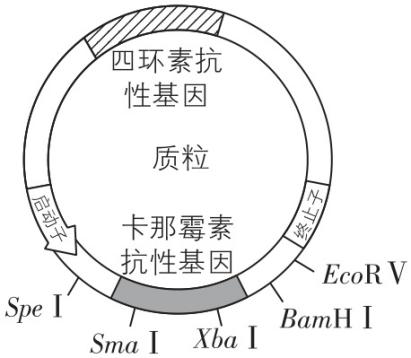
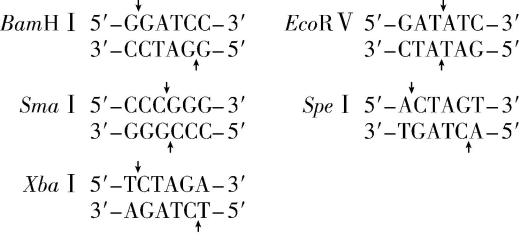
限制酶的识别序列和切割位点如下:
(3) 为将构建好的表达载体转入大肠杆菌,需要先用 $ {\mathrm{C}\mathrm{a}}^{2+} $ 处理大肠杆菌细胞,目的是 ,随后在含 和 的培养基中培养一段时间后,根据菌落周围有无水解透明圈筛选目的菌株。
答案:(1) 在红藻原生环境中筛选到具有卡拉胶降解能力的微生物的可能性大;稀释
(2) $ Bam\mathrm{H}Ⅰ $ ; $ Sma $ Ⅰ和 $ Eco\boldsymbol{R}Ⅴ $ 切割后产生平末端, $ \mathrm{S}pe $ Ⅰ和 $ Xba $ Ⅰ切割后产生的黏性末端相同,两种情况均会导致 $ cg $ 反向连接,新的序列无法被原有酶识别
(3) 使细胞处于一种能吸收周围环境中 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的生理状态;四环素;卡拉胶
解析:(1) 海洋红藻含卡拉胶,即海藻是卡拉胶的天然来源,而海泥中可能富集了海藻残体,因此,这两类样本中可能存在能高效降解卡拉胶的菌种。将培养后的菌液混匀并充分稀释,目的是使接种后的微孔板的一个孔中最多有一个菌种,确保后续筛选的菌种为纯培养物。
(2) $ \mathrm{P}\mathrm{C}\mathrm{R} $ 产物与质粒经双酶切产生的末端可用 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶连接,其中 $ E.coli\mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶连接具有平末端的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 片段的效率远低于 $ \mathrm{T}4\mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶。为保证连接准确性, $ cg $ 转录模板链的 $ 5\prime $ 端应位于重组质粒中靠近终止子的一端,根据质粒上限制酶的种类和酶切位点可知, $ cg $ 转录模板链的 $ 5\prime $ 端最好含有 $ Bam\mathrm{H}Ⅰ $ 的酶切位点。另两组同学选用了各不相同的双酶切组合并用 $ \mathrm{T}4\mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶连接,连接后形成的序列不再是原限制酶的识别序列,故无法使用各自构建表达载体时的双酶组合进行切割。
(3) $ {\mathrm{C}\mathrm{a}}^{2+} $ 处理可以提高细胞膜通透性,使细菌处于一种能吸收周围环境中 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的生理状态,便于外源 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 的导入。能表达 $ \mathrm{C}\mathrm{G} $ 的菌株可降解卡拉胶,在平板上形成透明圈,重组载体含四环素抗性基因,转化成功的菌株可在含四环素的培养基中存活,而卡那霉素抗性基因可能被破坏,不能作为筛选标记。